La photosynthèse est une des fonctions fondamentales du monde vivant. C'est cette fonction qui permet la croissance des plantes et donc la base de toute pyramide alimentaire et qui permet aussi la présence de l'oxygène sur Terre.
Le pivot du couple respiration/photosynthèse est le carbone.

En simplifiant à l'excès, on peut dire que le carbone minéral (dioxyde de carbone) est réduit en carbone organique (glucose) par la photosynthèse : on passe d'un carbone accompagné de 2 oxygènes à un carbone accompagné d'un oxygène. La respiration est donc une oxydation.
Document

L'expérience suivante a été faite les années précédentes. Elle n'est citée ici qu'en tant que rappel.


Feuille de Pelargonium après traitement

Feuille de Coleus à l'état naturel

Feuille de Coleus après traitement

Les résultats et les interprétations de cette expérience préliminaires sont donnés dans le tableau suivant :

Mise en évidence de la photosynthèse et structure du chloroplaste
Document

La pénétration du CO2 dans la plante peut être mise en évidence par une expérience d'autoradiographie.

Le carbone du CO2 est du 14C radioactif. La radioactivité permet de suivre le devenir de ce carbone. Il est intégré dans la feuille sous forme de glucose puis la sève élaborée va le transporter. C'est ce qui explique la présence de radioactivité dans la tige.
Que se passe t-il au niveau de la feuille ?
On observe au microscope une feuille d'Elodée du Canada, une plante aquatique très utilisée dans les bassins et les aquariums.
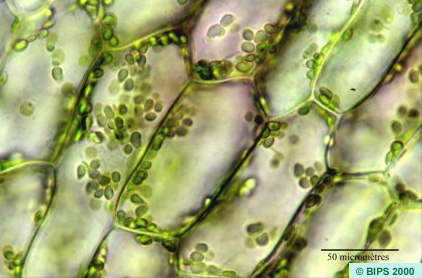
On observe facilement les chloroplastes. Si on laisse la préparation quelques minutes, on observe un déplacement des chloroplastes appelés mouvement de cyclose. Ce mouvement est beaucoup moins évident si on a une microscope éclairé par une ampoule LED, qui ne chauffe pas.
On peut tenter une coloration des chloroplastes par le lugol (eau iodée), mais franchement, je n'ai jamais eu de résultats très probants. Le schéma ci-dessous illustre l'expérience :

Les deux photos suivantes sont des électronographies (photos au microscope électronique) de chloroplastes.

On peut en tirer un schéma de la structure du chloroplaste permettant de mettre en évidence les structures chlorophylliennes.

Le rejet de dioxygène par des feuilles d'Elodée peut être mis en évidence par un montage ExAO. On utilise une solution d'hydrogénocarbonate de sodium de façon à ce que les feuilles reçoivent une grande quantité de CO2.

Si on a de la chance (un montage ExAO, ça marche rarement !), on peut obtenir une courbe de ce type :

Durant les phases d'obscurité, on constate une baisse du taux de dioxygène dans l'enceinte. Cette baisse met en évidence le phénomène de respiration des feuilles. Lorsque les feuilles sont éclairées, le taux de dioxygène augmente. La respiration continue, mais un autre phénomène vient la masquer, la photosynthèse qui entraîne une libération d'O2 par le feuilles.
Une autre manipulation (qui marche, celle-la) consiste à éclairer un rameau d'Elodée en éloignant progressivement celui-ci de la source de lumière.

On compte alors le nombre de bulles dégagées par la plante au niveau de la coupure de la tige. On constate que plus la source de lumière est proche et plus le nombre de bulles est élevé, c'est à dire que la photosynthèse est intense. Non seulement la lumière permet la photosynthèse, mais son intensité joue sur celle-ci.
Composition et propriétés lumineuses de la chlorophylle
Document

Une des techniques les plus simples et efficaces pour connaître la composition d'un mélange de substance est la chromatographie. C'est une chromatographie sur papier que nous allons utiliser pour connaître la composition de la chlorophylle. On récupère la chlorophylle directement sur la feuille.

On constate que la chlorophylle brute est formée de 4 pigments :
— chlorophylle b
— chlorophylle a
— xanthophylles
— carotènes
Lorsqu'on observe les formules chimiques, on constate que chlorophylle a et b sont très proches tout comme les carotènes et les xanthophylles qui appartiennent au groupe des caroténoïdes.
Formule chimique des chlorophylles

Formules chimiques des caroténoïdes de la chlorophylle brute

On peut aussi faire une séparation entre caroténoïdes et chlorophylles en ampoule à décanter en utilisant une solution alcoolique de chlorophylle brute, du dichloroéthane et de l'eau.

Le dichloroéthane est non miscible avec le mélange eau-alcool. On obtient donc deux phases, une phase supérieure (épiphase) jaune contenant les caroténoides et une phase inférieure (hypophase) verte contenant les deux chlorophylles.
Nous avons vu que la lumière et la chlorophylle étaient deux des acteurs majeurs de la photosynthèse. Quelles sont les relations entre les deux ?
Pour analyser le comportement de la chlorophylle en présence de lumière, on extrait d'abord de la chlorophylle de feuille d'épinard avec de l'éthanol.

A partir de cette solution, on peut d'abord analyser le spectre d'absorption de la chlorophylle. Deux méthodes sont utilisées : la spectroscopie et la spectrophotométrie.
Pour la spectroscopie, on utilise des petits spectroscopes à main (notre spectroscope historique donnant des signes de fatigue).

On obtient les résultats suivants :

On peut donc constater que la lumière dans les longueurs d'onde du bleu et d'une partie du rouge est absorbée.
Pour la spectrophotométrie, on utilise un spectrophotomètre couplé à un montage ExAO. On obtient directement le spectre d'absorption de la chlorophylle.
Un spectrophotomètre

Spectre d'absorption de la chlorophylle brute

Une autre caractéristique de la chlorophylle est sa réaction aux UV (365 nm).

La chlorophylle se colore en rouge foncé. C'est le phénomène de fluorescence.

Le schéma suivant donne une interprétation de ce phénomène qui n'apparaît que dans une solution de chlorophylle et jamais dans une feuille.

Lorsque un photon, correspondant à une longueur d'onde bleu (donc très énergétique) frappe un électron d'un atome de la chlorophylle, celui-ci est excité et change de niveau d'énergie. Il est donc instable. Il revient donc à son niveau d'énergie initiale en redonnant un photon. Comme un peu d'énergie a été perdu, le photon émis est moins énergétique et donc d'une longueur d'onde supérieur, c'est à dire dans le rouge. C'est ce qui explique la teinte prise par le chlorophylle.

Les relations entre les propriétés optiques de la chlorophylle et l'activité photosynthétique
Documents


On sait que les longueurs d'ondes absorbées par la chlorophylle correspondent aux couleurs rouges et bleus. On sait aussi que le photosynthèse se manifeste par un rejet de dioxygène. Afin de relier les deux observations, on va tester l'efficacité d'un éclairement en vert, bleu et rouge sur la photosynthèse.
Deux techniques sont utilisables :
— un montage ExAO permettant de mesurer le taux d'oxygène de l'enceinte en fonction d'un éclairement donné par une ampoule LED à couleurs variables.

— un comptage de bulle sur un rameau d'élodée en fonction de la couleur de l'éclairement donné par une ampoule LED.
Les résultats obtenus sont très variables et parfois difficilement exploitables. On peut obtenir une courbe de ce type (le bleu donne des résultats non exploitables).

On peut donc comparer le spectre d'action (rejet de dioxygène) de la chlorophylle brute à son spectre d'absorption (ici on a séparé les différents pigments).

On constate qu'il y a une bonne correspondance entre les deux types de spectres.
Comment expliquer alors la relation entre rejet de dioxygène et rôle de la lumière ?
En 1941, Hill propose l'expérience suivante : il fait un broyat de feuilles qu'il introduit dans une enceinte en présence d'un oxydant puissant (appelé réactif de Hill). Dans cette version de l'expérience de Hill, l'oxydant est du DCPIP.

On constate que lorsque la suspension est éclairé et en présence de DCPIP, il y a libération de dioxygène. Nous avons vu que lorsque la chlorophylle est éclairée, des électrons sont excités. On sait qu'un oxydant est avide d'électrons. Les électrons excités passent donc de la chlorophylle au DCPIP.
Deux questions se posent alors :
— comment la chlorophylle récupèrent les électrons qui lui sont prélevés ?
— comment expliquer le changement de couleur du DCPIP ?
Au moment où Hill élaborait son expérience, deux autres chercheurs étudiaient la photosynthèse d'une autre manière. En effet, Ruben et Kamen cherchent à prouver que le dioxygène rejeté par la plante à pour origine le dioxygène contenu dans le CO2.

On constate que le dioxygène du CO2, radioactif, ne se retrouve pas dans l'oxygène rejeté par la plante. Il est intégré au glucose synthétisé. La seule source de dioxygène utilisable alors est donc l'eau. Or on peut écrire :

Les électrons perdus par la chlorophylle sont donc récupérés par la cassure de la molécule d'eau (photolyse de l'eau). En réalité ce sont bien des H+ et des électrons, soit des H2, que récupèrent la chlorophylle. Cela signifie que lors de l'expérience de Hill, ce sont des H2 qui sont transmis au DCPIP. On explique ainsi facilement le changement de couleur, du à la réduction de l'oxydant par les hydrogènes.
Les électrons étant plus énergétiques du fait l'absorption d'un photon, cette énergie va être stockée sous une forme chimique dans la molécule d'ATP (voir plus loin). Le transfert d'hydrogène se couple à une conversion de l'énergie lumineuse en énergie chimique. On appellera R, l'accepteur d'hydrogène dans la feuille, qui joue le rôle de l'oxydant de l'expérience de Hill.
Schéma simplifié de la phase photochimique de la photosynthèse

Interprétation d'expériences anciennes, les expériences d'Engelman

L'interprétation de ces expériences est très simple. On sait que Bacterium thermo est avide de dioxygène. Lorsque le filament d'algue est faiblement éclairée, l'intensité photosynthétique est très faible : il y a donc un faible rejet de dioxygène. B. thermo reste donc autour d'une bulle d'air. Lorsque l'éclairement est plus intense, le rejet de dioxygène par photosynthèse attire les bactéries.
La dernière expérience est une application du spectre d'action. Ce sont les rayonnements rouge et bleu qui sont actifs sur la chlorophylle, donc sur le rejet d'O2. L'accumulation de bactéries dans ces zones est alors facilement compréhensible.
La molécule d'ATP
La molécule d'ATP (adénosine triphosphate) est une forme de stockage temporaire de l'énergie dans la cellule. L'énergie cellulaire ne peut être utilisée que sous forme d'ATP.

Le stockage maximal d'énergie se fait au niveau de la liaison entre le second et le troisième phosphate. La réaction de stockage d'énergie par ajout d'un phosphate s'appelle une phosphorylation. Lorsque le dernier groupe phosphate est éliminé par hydrolyse, on parle alors de déphosphorylation.

Comme toute réaction chimique dans une cellule vivante, ces deux réactions sont catalysées par des enzymes, l'ATPase et l'ATPsynthase.

La phase non-photochimique ou phase d'assimilation
Documents


C'est pendant cette phase que le dioxyde de carbone est intégré dans une suite de réactions chimiques qui aboutit à la formation de molécules organiques (glucose).

Deux expériences préliminaires, ont précisées les conditions d'activités de cette phase d'assimilation : les expériences de Gaffron et d'Arnon.

L'expérience de Gaffron met en évidence que la fixation de CO2 ne nécessite pas de lumière. SI tel avait été le cas, le passage à l'obscurité aurait obligatoirement entraîné une arrêt brutal de la fixation de CO2, ce qui n'est pas le cas.
L'expérience d'Arnon montre que la fixation du CO2 se produit au niveau du stroma du chloroplaste.
La phase photochimique a lieu au niveau des tylakoïdes alors que la phase d'assimilation se passe au niveau du stroma.
Ce sont surtout les expériences de Calvin et ses collaborateurs qui ont permis de comprendre le mécanisme de la phase d'assimilation du CO2.

Durant cette expérience historiques, Calvin et al. utilisent des algues vertes microscopiques, les chlorelles. Pendant la phase d'assimilation, ils marquent le CO2 incorporé au 14C puis font des autoradiographies à des temps très proches. Ils peuvent ainsi suivre l'incorporation du carbone marqué dans les différentes molécules organiques. Ils constatent très vite que le pivot des réactions est marqué par la présence de deux molécules, le Ru-dP (ribulose di-phosphate) et l'APG (acide phosphoglycérique).

L'analyse de la première expérience montre que lorsqu'on bloque la phase photochimique par l'obscurité on a une baisse du Ru-dP et en même temps une augmentation du taux d'APG. Cela signifie que le Ru-dP est transformé en APG. Lorsque tout le Ru-dP a été consommé, le taux d'APG diminue puisqu'il est à son tour, intégré dans la suite des réactions biochimiques.
La seconde expérience montre que le CO2 est intégré entre le stade Ru-dP et le stade APG. Lorsque le taux de CO2 est nul, l'APG ne se forme plus mais le Ru-dP continue à se former durant un certain temps. Cela signifie que c'est l'APG qui conditionne la présence de Ru-dP.
Si on additionne les résultats des deux expériences, on est obligé de conclure qu'il existe un cycle fonctionnant entre le Ru-dP et l'APG. On appelle ce cycle, le cycle de Calvin-Benson ou plus simplement cycle de Calvin.

On constate que l'ATP récupéré lors de la phase photochimique est l'énergie servant à faire tourner le cycle et que les RH2 (accepteurs d'hydrogène réduits) sont régénérés en R (accepteurs oxydés) en cédant leurs hydrogènes au cycle. A chaque tour de cycle, un sucre en C3 (triose) est fabriqué et sorti du cycle. Il faut donc deux tours pour fabriquer une molécule de glucose.
La RubisCO est une enzyme fondamentale du monde végétal.

Cette enzyme intervient donc au moment où le CO2 se fixe, c'est à dire entre le stade Ru-dP et le stade APG.
Schéma général simplifié de le photosynthèse

C'est très important
RépondreSupprimermerci beaucoup
J'ai trouvé l'article tres interressant
RépondreSupprimertrès intéressant
RépondreSupprimerTrès intéressant
RépondreSupprimer